
 分享
分享

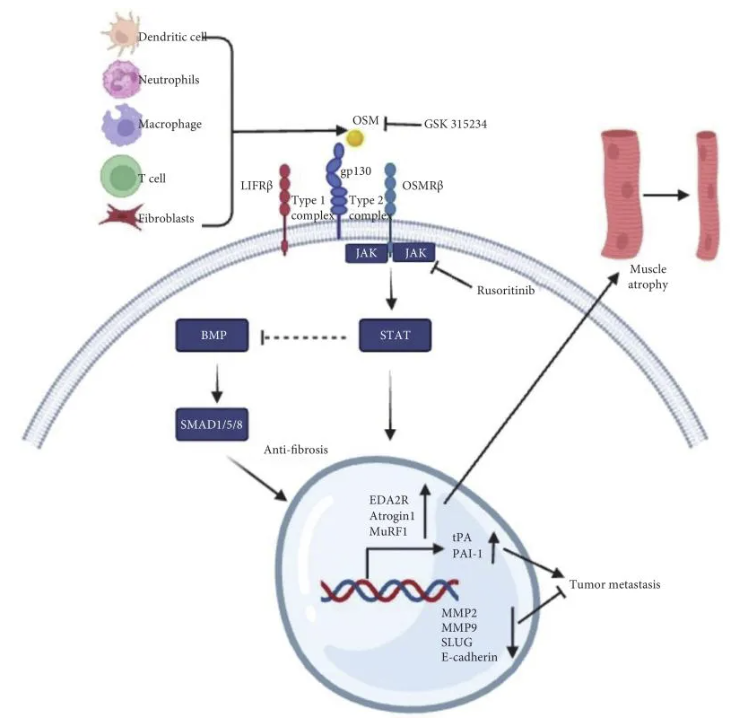
白细胞介素-6(Interleukin-6,IL-6)家族成员包括IL-6,抑瘤素M(oncostatin M,OSM),白血病抑制因子(Leukemia Inhibitory Factor,LIF),IL-11,IL-27,心肌营养素-1(cardiotrophin-1,CT-1)和心肌营养素样细胞因子1(cardiotrophin-like cytokine factor 1,CLCF1)参与调节各种炎症和肿瘤[1-3]。1986年,Zarling等[4]首次从人淋巴瘤U937细胞中分离一种生长调节蛋白,因其具有抑制黑色素瘤细胞的细胞生长而影响成纤维细胞因此得名OSM。成熟的人OSM分子大小约22 kDa,由196个氨基酸组成的可溶性蛋白[5]。OSM主要由活化的巨噬细胞、肿瘤相关成纤维细胞(cancer-associated fibroblasts,CAF)、T细胞、树突状细胞和中性粒细胞等多种细胞合成和分泌(图1)[1,5]。LIF和OSM在结构上是IL-6家族最相似的成员,与其他IL-6家族细胞因子成员类似,OSM与gp130细胞外细胞因子结合同源区(cytokine-binding homology region,CHR)结构域结合,随后招募LIFRβ形成I型受体复合物(LIFRβ/gp 130)或OSMR β形成II型受体复合物(OSMRβ/gp 130),且OSM与LIFR的结合亲和力明显低于OSMR(II型)复合物,激活Janus活化激酶/信号转导子和转录激活子3(Janus-activated kinase/signal transducer and activator of transcription 3,JAK/STAT 3)、促分裂原活化蛋白激酶/细胞外调节激酶(mitogen-activated protein kinase/extracellular regulator kinase,MAPK/ERK)、c-Jun N末端激酶(c-Jun N-terminal Kinase,JNK)和磷脂酰肌醇-3-激酶/蛋白激酶B(phosphatidylinositol-3-kinase/protein kinase B,PI3K/AKT)等信号通路参与多种生理和病理功能,调控炎症、肿瘤和肌肉萎缩等(图1)[6-10]。对于部分细胞类型,OSM信号传导亦可激活STAT1和STAT5通路[11-12]。目前OSM与OSMR形成的II型复合物通路在各种细胞系及动物实验中进行了广泛研究,LIFR(I型)复合物如何影响信号传导或疾病进展尚不明确[1]。OSM已被证实在肺部炎症和肿瘤中发挥重要作用,然而抗OSM治疗目前尚在临床前研究。本文综述OSM与慢性肺部疾病的关联, 介绍抗OSM通路对慢性肺部病变和肺部肿瘤的研究进展。
1、OSM与慢性气道疾病
细胞因子OSM在慢性气道疾病过程中参与细胞外基质(Extracellular Matrix,ECM)蛋白沉积(如肺纤维化或严重哮喘)或分解代谢(如慢性阻塞性肺疾病)调控、在细胞增殖和细胞存活等方面发挥关键作用。肺成纤维细胞募集到气道损伤部位被认为是导致ECM过度沉积的重要原因,且转化生长因子-β(transforming growth factor-β,TGF-β)已被证实是纤维化发病机制的重要调节因子,TGF-β非依赖性途径也可能参与其中[13]。
1.1 肺纤维化
过度的成纤维细胞活化、积聚和表型变化可能导致肺纤维化和组织功能丧失重要原因。Mozaffarian等[14]首次发现OSM在特发性肺纤维化和硬皮病患者的支气管肺泡灌洗液中表达增高,募集炎症细胞促进胶原沉积增加,该效应不依赖B和T淋巴细胞、嗜酸性粒细胞和肥大细胞且不通过经典IL-4/IL-13和TGF-β途径,提示OSM可能作为独立因素参与特发性肺纤维化的进程。Esnault[15]等通过独创性途径分析和qPCR进一步验证表明OSM是成纤维细胞激活的潜在上游调节因子。过量的STAT-1/STAT 3信号传导显著加剧博莱霉素诱导的小鼠肺纤维化,而不通过经典的TGFβ-SMAD 3途径[16]。因此,OSM通过何种机制诱导肺纤维化则成为研究的热点。在BALB/c小鼠肺中,OSM诱导显著的pSTAT 3活化,伴随pSMAD 1/5抑制,且无TGFβ -SMAD 2活化[17]。由于pSMAD 1/5/8是BMP信号通路的下游,其在肺纤维化模型中表现出保护作用[18],因此pSTAT 3的过度活化可能通过调节BMP/pSMAD 1途径间接介导纤维化。Ayaub等[19]通过博来霉素诱导的肺纤维化模型,并经过腺病毒转染高表达IL-6和OSM,结果表明暴露于博来霉素和OSM或IL-6的小鼠肺中M2样巨噬细胞的数量显著增加,高表达IL-6Rα且缺乏OSMRβ,提出IL-6和OSM参与诱导巨噬细胞的选择性M2活化,恶化博来霉素诱导的肺纤维化。进一步研究发现OSM增强成纤维细胞趋化性是由信号转导子和转录激活子(STAT3)和p38丝裂原活化蛋白激酶介导的,而凝胶收缩和α-SMA表达的增强则由STAT3介导的[20]。与既往研究不同,Lan等[21]发现OSM预处理的间充质干细胞经旁分泌肝细胞生长因子,促进细胞增殖和迁移,博来霉素诱导的肺纤维化小鼠中移植OSM预处理的充质干细胞可显著改善肺的呼吸功能,并下调了肺组织中炎症因子转化生长因子-β1和纤维化因子的表达。然而,OSM在特发性肺纤维化的作用仍需进一步阐述明确。
1.2 哮喘和慢性阻塞性肺疾病
OSM参与气道重塑和肺功能下降相关,重症哮喘由于气道重塑致气流不完全可逆,因此OSM可能调控重症哮喘和慢性阻塞性肺疾病的病理改变。Simpsont等[22]发现相比于控制稳定不吸烟的哮喘患者,重症哮喘伴不完全可逆气流受限患者的痰液中巨噬细胞和中性粒细胞OSM表达水平明显上升。近期经单细胞测序技术进一步证实气道内OSM由肺炎克雷伯杆菌的脂多糖激活的巨噬细胞分泌[23]。Baines等[24]亦发现相比于健康志愿者,COPD患者的诱导痰中IL-8 和OSM表达明显增高,提示OSM与重症哮喘和慢性阻塞性肺疾病的慢性气道炎症和气道重塑有关。Lin等[25]经生物信息学算法与单细胞或转录组数据进行交叉分析结果表明高水平的OSM,IL-18R1与重症哮喘患者的肺功能和生存时间成负相关,因此OSM可作为重症哮喘的监测和预后的标志物。在严重哮喘中,气道上皮粘液产生细胞数量增加、肺间质纤维化及巨噬细胞高表达卵泡抑素样1促进OSM过度表达,诱导气道上皮细胞中抵抗素样分子α的增加,导致小鼠肺细胞外基质重塑[26],而抗OSM单抗可逆转卵泡抑素样1对气道的重塑,提示OSM途径可能是作用重症哮喘和慢性阻塞性肺疾病气道重塑新的靶点。
2、OSM与肺癌
肺癌是发病率和死亡率最高的恶性肿瘤,分子靶向和免疫的治疗改善患者的生存时间,但是,多数患者在治疗后仍死于肿瘤的耐药、复发 [27],因此迫切需要寻找高效低毒新的治疗靶点。OSM最初被发现在体外对黑色素瘤和其他癌症细胞系具有直接的增殖抑制作用[4],Ouyang[28]等发现低浓度OSM在体外可抑制人肺癌细胞95-D细胞的增殖,减少基质金属蛋白酶2(matrix metalloproteinase-2,MMP-2)和MMP-9的分泌降低细胞的粘附和侵袭能力。Spence等[29]发现OSM通过JAK3/STAT3通路促进组织型纤溶酶原激活剂(type plasminogen activator,tPA)和纤溶酶原激活物抑制物-1(plasminogen activator inhibitor-1,PAI-1)mRNA的表达促进肿瘤转移。McCormick和Cichy团队发现OSM较IL-6可更有效的促进肺癌细胞分化,OSM与TGF β 1因子共同调节透明质酸,促进肺癌转移[30-31]。与之前的研究不同,Lauber等[32]发现OSM在体外对LLC细胞无增殖抑制作用,但在小鼠体内可促进肿瘤细胞显著增长,通过转染腺病毒致肺部OSM表达增高,可募集M2巨噬细胞改变肿瘤微环境,促进异位种植的肿瘤肺转移增加,然而OSMRβ 敲除鼠则减少肿瘤肺转移,提示OSM/OSMRβ通路调控肿瘤肺转移。Shien等[33]亦发现OSM可在肺癌小鼠模型中经上皮间质转化(epithelial–mesenchymal transition,EMT)促进肺转移,肺癌细胞与CAF体外共孵育后,磷酸化STAT 3、OSMRβ和LIFRβ上调,同时E-钙粘蛋白下调,CAF经OSMRβ/JAK 1/STAT 3通路促进肿瘤进展和耐药,靶向选择性JAK1抑制剂非戈替尼有效抑制STAT3激活,降低OSMR表达,抑制肿瘤增殖逆转肿瘤耐药。临床前研究发现OSM及其通路在肺癌的发生和转移中起到关键作用,那么OSM在真实世界肺癌患者中扮演什么样的角色?Shien等[33]及其团队分析TGCA和PROSPECT公共数据库,高表达OSM,IL-6,LIF和OSMRβ的肺腺癌患者预后更差。另有研究表明放疗和缺氧应激的肺癌细胞分泌大量微泡,诱导CAF分泌包括OSM,IL-8,IL-11,血管内皮生长因子等[34],OSM可能参与肿瘤的自我适应调控。部分研究发现OSM通过STAT1信号通路下调EMT起动子SLUG转录,抑制EMT及肺癌转移[35]。综上,临床研究表明OSM可促进肿瘤增殖和转移,而临床前研究的结果并不完全一致,OSM通路网络在肺癌中的具体机制需要进一步研究阐明。
3、OSM与恶病质
恶病质是一种以消耗为特征复杂的多因素综合征,在肺癌和慢性肺部疾病患者中普遍存在,其主要特征表现为体重减轻、骨骼肌质量减少(伴或不伴有脂肪组织损失)以及全身炎症[36-38]。全身性炎症因子包括肿瘤坏死因子-α(TNF-α)、白细胞介素-1(IL-1)和IL-6 等水平升高可导致肌肉萎缩,针对其相应的中和抗体可抑制其萎缩效应[39-40]。在胰腺导管腺癌(高达80%患者发生恶病质)和肌营养不良患者的肌肉中OSMR和OSM靶基因的转录水平显著增加,提示其在恶病质相关的肌肉萎缩中发挥重要作用[41]。Miki等[42]发现OSM处理C2C12肌管细胞24 h和48 h后,肌细胞生成素表达下降,atrogin-1和OMSR表达上升,肌管直径比对照组分别减少18.7%和23.3%,STAT3抑制剂或敲除STAT3基因可逆转其效应。与之类似,Domaniku等[41]通过敲除荷瘤小鼠特异性肌肉细胞OSMR,可抑制肌肉细胞萎缩恢复肌肉功能,近期其团队进一步证实OSM可在体外通过JAK/STAT3通路上调原代肌管细胞表达Atrogin1致肌肉萎缩,明确了OSM信号通路在肌肉萎缩中的作用[43]。OSM诱导的肌肉萎缩依赖于JAK/STAT 3信号传导,与Atrogin 1、Ampd 3、Serpina 3n、Cebpd、Sln和Mt 1/2多种萎缩相关基因的表达增加有关[42]。Bilgic等[43]近期工作发表在Nature杂志首次发现肺癌患者和Lewis小鼠恶病质模型的肌肉中组织外胚层发育不良A2受体(ectodysplasin A2 receptor,EDA2R)表达异常升高,与EDA2结合激活非经典NF-ĸB信号通路促进Atrogin1和MuRF1基因转录和蛋白表达,从而诱导肌肉萎缩,有趣的是,OSM不仅与OSMR结合发挥作用,可同时促进EDA-2R受体转录表达加剧肌肉细胞萎缩,敲除肌肉OSMR受体可逆转EDA2R受体表达,抑制恶病质相关肌肉萎缩基因转录和缓解肌肉功能受损。相比与IL-6和LIF,OSM对萎缩基因的转录的调控作用更强,因此对肌管的萎缩效应更明显。由于这三种细胞因子都与肌肉萎缩有关,因此理论上针对这些途径的共同通路可能更有效。因此,近期一项I期针对IV期非小细胞肺癌合并恶病质的患者中使用JAK 1/2抑制剂鲁索利替尼的安全性和有效性的临床试验(NCT 04906746)正在募集患者。一项II期随机对照试验,针对人源化单克隆抗OSM抗体(GSK 315234)和安慰剂在类风湿性关节炎的疗效研究,虽未达到其预定的研究终点,初步表明人源性OSM中和抗体在人体内使用的安全性和耐受性[44]。此外,中和性抗OSM抗体在Lewis小鼠恶病质模型中已显示出改善肌肉萎缩和肌肉功能。开发针对OSM信号通路的靶向治疗药物对于治疗包括肺癌在内的多种疾病至关重要,期待后续更多针对OSM通路的药物的研发为临床治疗提供新的希望。
OSM在多种肺部疾病中发挥重要作用,部分抗OSM通路药物临床前研究取得优异的疗效,然而目前尚无药物获批用于临床疾病的治疗。未来针对抗OSM或抗OSMRβ的药物治疗将成为肺部疾病研究中的热门领域,我们期待着后续针对OSM信号通路更加深入的研究以及更多有效安全药物的出现并应用于临床。
利益冲突:本研究不涉及任何利益冲突。
参考文献(略)
作者介绍

第一作者:柯章敏
南京市江宁医院呼吸与危重症医学科副主任医师(在职博士);南京市五一劳动奖章获得者;获实用新型专利二项;获南京市医学会青年科技奖一项;主持及参与国自然及市级课题三项,发表SCI及医学核心期刊论文10余篇。

通讯作者:宋勇
教授;南京大学医学院附属金陵医院呼吸与危重症医学科主任;南京大学呼吸病学研究所所长;南京大学、南方医科大学、南京医科大学、东南大学博士研究生导师;江苏省医学会呼吸病分会主任委员;中华医学会呼吸分会全国委员;南京医学会呼吸分会前任主任委员;《Translational Lung Cancer Research》杂志(SCI收录)主编。
引用本文:柯章敏, 潘航程, 王丽, 吕镗烽, 张秀伟, 宋勇. 抑瘤素M在肺部疾病及恶病质中的研究进展. 中国呼吸与危重监护杂志, 2024, 23(12): 900-904. doi: 10.7507/1671-6205.202401039
本文转载自「中国呼吸与危重监护杂志」
* 文章仅供医疗卫生相关从业者阅读参考
本文完
责编:Jerry